

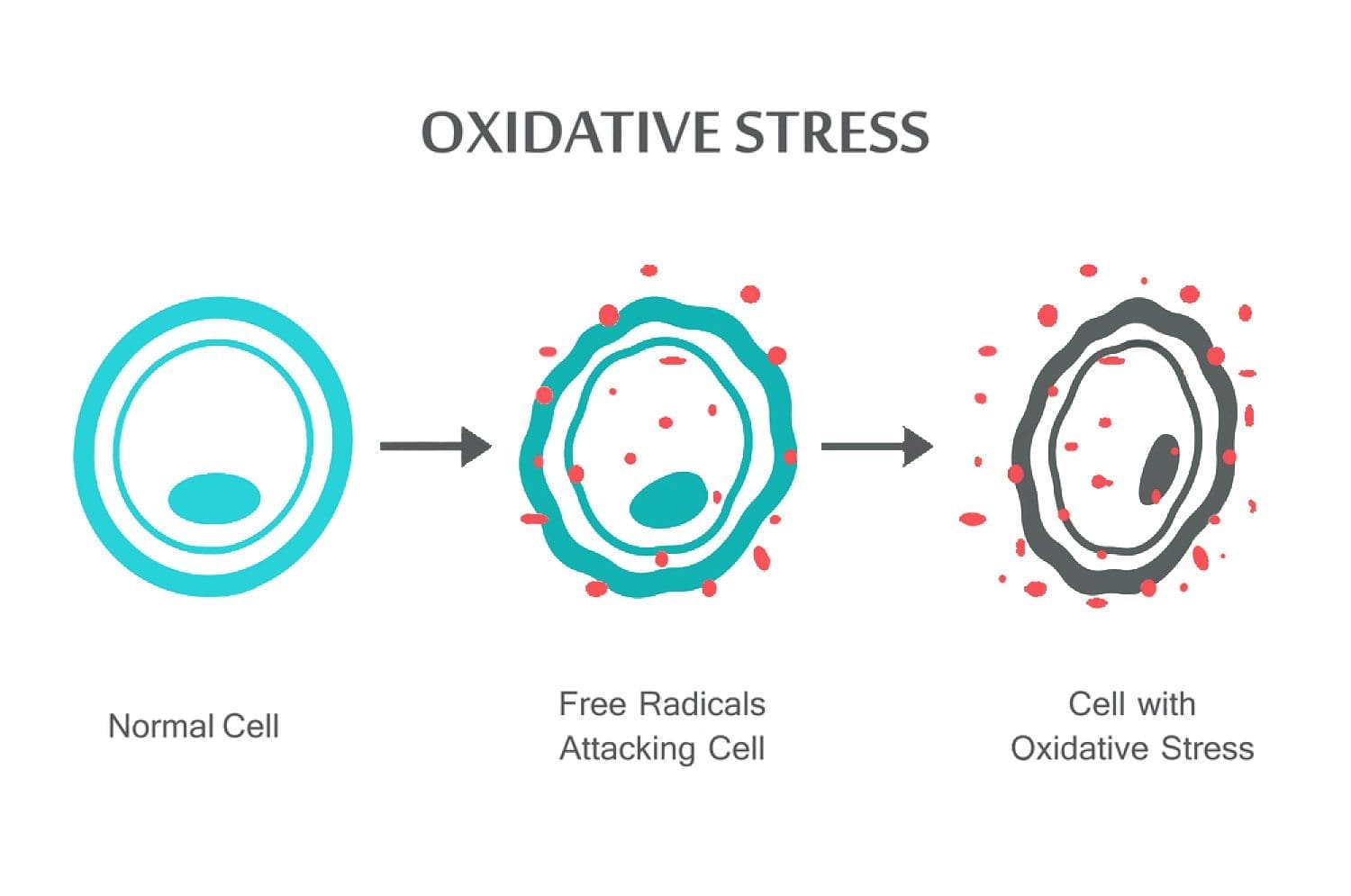
Vísindamiðað Chiropractor Dr. Alexander Jimenez kíkir á oxun streitu, hvað það er, hvernig það hefur áhrif á líkamann og andoxunarefni vörn til að ráða bót á ástandinu.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD, 2 og Omer Kalayci, MD1
Samantekt: Reactive oxygen species (ROS) eru framleiddar af lifandi lífverum vegna eðlilegrar umbrots og umhverfisþátta, svo sem loftmengunarefna eða sígarettureykja. ROS eru mjög hvarfgjarn sameindir og geta skaðað frumustofnanir eins og kolvetni, kjarnsýrur, fituefni og prótein og breytt virkni þeirra. Vöktunin í jafnvægi milli oxunarefna og andoxunarefna í þágu oxandi efna er nefnt "oxandi streita." Reglugerð um að draga úr og oxandi (redox) ástandi er mikilvægt fyrir lífvænleika, virkjun, útbreiðslu og líffæravirkni. Þolfimi lífverur hafa samþætt andoxunarefni kerfi, sem fela í sér ensím og ekki ensímandi andoxunarefnum sem eru yfirleitt árangursríkar við að hindra skaðleg áhrif ROS. Hins vegar, við sjúkdómsástand, geta andoxunarefnin verið óvart. Oxidandi streita stuðlar að mörgum sjúkdómsástandi og sjúkdómum, þar með talið krabbamein, taugasjúkdóma, æðakölkun, háþrýstingur, blóðþurrð / ofsakláði, sykursýki, bráða öndunarfærasjúkdómur, lungnasjúkdómur í lungum, langvinna lungnateppu og astma. Í þessari umfjöllun er samantekt á frumueyðandi og andoxunarefnum og fjallað um frumuáhrif og aðferðir oxunarálagsins.
Lykilorð: andoxunarefni, oxunarefna, oxandi streita, hvarfefni súrefnis, redox
(WAO Journal 2012; 5: 9-19)
Viðbrögð súrefnistegunda (ROS) eru framleidd af lifandi lífverum vegna eðlilegrar efnaskipta frumna. Við lágan til í meðallagi mikinn styrk virka þeir í lífeðlisfræðilegum frumuferlum, en við háan styrk mynda þeir skaðlegar breytingar á frumuhlutum, svo sem lípíðum, próteinum og DNA.1–6 Breytingin á jafnvægi milli oxunar / andoxunarefnis í þágu oxunarefna er kallað „oxunarálag“. Oxunarálag stuðlar að mörgum sjúklegum aðstæðum, þar á meðal krabbameini, taugasjúkdómum, 7–10 æðakölkun, háþrýstingi, blóðþurrð / perfusion, 11–14 sykursýki, bráðri öndunarerfiðleikarheilkenni, sjálfvakinni lungnatrefju, langvarandi lungnateppu, 15 og astma.16– 21 Loftháðar lífverur hafa samþætt andoxunarefni kerfi, sem fela í sér ensím og ó ensímfræðileg andoxunarefni sem eru venjulega áhrifarík til að hindra skaðleg áhrif ROS. Hins vegar, við sjúklegar aðstæður, geta andoxunarefni kerfin verið óvart. Í þessari endurskoðun erum við að draga saman frumuoxíðandi og andoxunarkerfi og stjórna afoxandi og oxandi (redox) ástandi í heilsufars- og sjúkdómsástandi.
Efnisyfirlit
Oxunarefni
Innrænar heimildir ROS
ROS eru framleidd úr sameinda súrefni vegna eðlilegrar umbrotsefna í frumum. ROS má skipta í 2 hópa: sindurefna og óradískur. Sameindir sem innihalda einn eða fleiri ópaired rafeindir og þannig gefa viðbrögð við sameindinni eru kölluð sindurefna. Þegar 2 sindurefnum deilir ópöruðu rafeindunum eru óradískar formir búnar til. 3 meiriháttar ROS sem eru af lífeðlisfræðilegu þýðingu eru superoxíðanjón (O22.), Hýdroxýlstakeind (OH) og vetnisperoxíð (H2O2). ROS eru teknar saman í töflu 1.
 Súperoxíð anjón myndast með því að bæta 1 rafeind við sameindasúrefnið.22 Þetta ferli er miðlað af nikótín adenín dínukleótíð fosfati [NAD (P) H] oxidasa eða xantín oxidasa eða með hvatbera rafeindaflutningskerfi. Helsta staðurinn til að framleiða súperoxíðanjón er hvatberar, vélar frumunnar til að framleiða adenósín þrífosfat. Venjulega eru rafeindir fluttar um rafeindaflutningskeðju hvatbera til að draga úr súrefni í vatn, en um það bil 1 til 3% allra rafeinda leka úr kerfinu og framleiða súperoxíð. NAD (P) H oxidasa er að finna í fjölfrumukjarna hvítfrumum, einfrumum og stórfrumum. Við phagocytosis framleiða þessar frumur sprengju af súperoxíði sem leiðir til bakteríudrepandi virkni. Súperoxíði er breytt í vetnisperoxíð með virkni súperoxíð dismutasa (SODs, EC 1.15.1.1). Vetnisperoxíð dreifist auðveldlega yfir plasmahimnuna. Vetnisperoxíð er einnig framleitt með xantínoxidasa, amínósýruoxidasa og NAD (P) H-oxidasa 23,24 og í peroxísómum með því að neyta sameindasúrefnis í efnaskiptum. Í röð viðbragða sem kallast Haber – Weiss og Fenton viðbrögð getur H2O2 brotnað niður í OH2 í viðurvist smitmálma eins og Fe21 eða Cu21.25
Súperoxíð anjón myndast með því að bæta 1 rafeind við sameindasúrefnið.22 Þetta ferli er miðlað af nikótín adenín dínukleótíð fosfati [NAD (P) H] oxidasa eða xantín oxidasa eða með hvatbera rafeindaflutningskerfi. Helsta staðurinn til að framleiða súperoxíðanjón er hvatberar, vélar frumunnar til að framleiða adenósín þrífosfat. Venjulega eru rafeindir fluttar um rafeindaflutningskeðju hvatbera til að draga úr súrefni í vatn, en um það bil 1 til 3% allra rafeinda leka úr kerfinu og framleiða súperoxíð. NAD (P) H oxidasa er að finna í fjölfrumukjarna hvítfrumum, einfrumum og stórfrumum. Við phagocytosis framleiða þessar frumur sprengju af súperoxíði sem leiðir til bakteríudrepandi virkni. Súperoxíði er breytt í vetnisperoxíð með virkni súperoxíð dismutasa (SODs, EC 1.15.1.1). Vetnisperoxíð dreifist auðveldlega yfir plasmahimnuna. Vetnisperoxíð er einnig framleitt með xantínoxidasa, amínósýruoxidasa og NAD (P) H-oxidasa 23,24 og í peroxísómum með því að neyta sameindasúrefnis í efnaskiptum. Í röð viðbragða sem kallast Haber – Weiss og Fenton viðbrögð getur H2O2 brotnað niður í OH2 í viðurvist smitmálma eins og Fe21 eða Cu21.25
Fe31 + .O2? Fe2 + O2 Haber Weiss
Fe2 + H2O2? Fe3 + OH + .OH Fenton viðbrögð
O 2 sjálft getur einnig hvarfast við H2 og myndað OH .2 Hýdroxýlhópur er mest hvarfgjarn af ROS og getur skaðað prótein, lípíð og kolvetni og DNA. Það getur einnig byrjað fituofoxun með því að taka rafeind úr fjölómettuðum fitusýrum.
Granulocytic ensím auka enn frekar virkni H2O2 með eosinophil peroxidasa og myeloperoxidasa (MPO). Í virkum daufkyrningum er H2O2 neytt af MPO. Í nærveru klóríðjóns er H2O2 breytt í hypochlorous sýru (HOCl). HOCl er mjög oxandi og gegnir mikilvægu hlutverki við að drepa sýkla í öndunarvegi. 28 HOCl getur einnig bregst við DNA og örva DNA-próteinskipti og framleiða pyrimidínoxunarefni og bæta klóríði við DNA basa. 29,30 Eosinophil peroxidasa og MPO stuðla einnig að oxunarálagi með breytingu á próteinum með halógeneringum, nítrunar- og próteinhleðsluskilum með tyrosýlstakeindum.31-33
Aðrar súrefnistengdu sindurefnum eru peroxýlstakeindirnar (ROO $). Einfaldasta form þessara róttækja er hýdroxperoxýlstakeind (HOO $) og hefur hlutverk í fitusýruperoxíðun. Frívalsfrumur geta kallað á lípíðperoxidation keðjuverkanir með því að draga úr vetnisatóm úr hliðarkeðju metýlen kolefni. Fituhjúpurinn hvarfast síðan við súrefni til að framleiða peroxýlstakeind. Peroxýlstakefni hefst keðjuverkun og umbreytir fjölómettaðar fitusýrur í lípíðhýdroxperoxíð. Lipíðhýdroperoxíð eru mjög óstöðug og geta auðveldlega sundrast í efri vörur, svo sem aldehýð (svo sem 4-hydroxy-2,3-nonenal) og malondialdehýð (MDA). Ísóprostir eru önnur hópur af fituefnumoxunarefnum sem myndast í gegnum peroxidation arakidonsýru og hefur einnig reynst hækka í plasma og öndunarsamdrætti astmatískra manna. 34,35 Peroxíðun lípíða truflar heilleika frumuhimna og leiðir til endurskipulagningar á himnauppbyggingu .
Vatnperoxíð, superoxíðstakefni, oxað glútaþíon (GSSG), MDAs, ísópróstan, karbónýl og nítrotýrosín geta hæglega verið metnar úr plasma-, blóð- eða berkjalokumolunarsýnum sem lífmælingar á oxun með stöðluðu prófunum.
Exogenous Source of Oxidants
Sígarettu Reykur
Sígarettureykur inniheldur margar oxandi efni og sindurefna og lífræna efnasambönd, svo sem súrefni og köfnunarefnisoxíð. 36 Að auki bætir innöndun sígarettureyks í lunguna einnig nokkrar innrænar aðferðir, svo sem uppsöfnun daufkyrninga og stórfrumna, sem eykur enn frekar oxunareinkenni .
Ósonlosur
Ozone váhrif geta valdið losun peroxíða og valdið innrennsli daufkyrninga í öndunarvegiþekju. Skammvinn útsetning fyrir óson veldur einnig losun bólgueyðandi lyfja, svo sem MPO, katjónísk eósínfíkla prótein og einnig laktat dehýdrógenasa og albúmín. 37 Jafnvel hjá heilbrigðum einstaklingum veldur útsetning á völdum lungnastarfsemi. 38 Cho et al39 hefur sýnt að agnir (blanda af agnir og fljótandi dropar sem eru svifaðir í loftinu) hvetur súrefnislækkunina.
Ofnæmi
Ofnæmi vísar til aðstæðna hærri súrefnisþéttni en venjulegt hluta þrýstings á súrefni í lungum eða öðrum líkamsvefjum. Það leiðir til meiri framleiðslu á hvarfefnum súrefnis og köfnunarefnis. 40,41
Ionizing geislun
Jónandi geislun, í nærveru O2, breytir hýdroxýlstakefni, superoxíði og lífrænum stakeindum í vetnisperoxíð og lífrænar vatnsperoxíð. Þessar vetnisperoxíðategundir bregðast við rauðoxíðvirkum málmjónum, svo sem Fe og Cu, með Fenton-viðbrögðum og þannig stuðla að oxunarálagi. 42,43 Narayanan og al44 sýndu að fibroblasts sem voru útsett fyrir alfa agnir höfðu verulega aukningu á innanfrumu O2 2. og H2O2 framleiðsla með plasmafrumumbundnu NADPH oxidase.44 Signal transduction sameindunum, svo sem utanfrumu-merki-reglubundnu kínasa 1 og 2 (ERK1 / 2), C-Jun N-endanlega kínasa (JNK) og p38 og umbrotsefni, eins og virkjunarprótein-1 (AP-1), kjarnorkuþáttur-kB (NF-kB) og p53, eru virkjaðir, sem leiða til tjáningar á geislavirkni sem tengist geislum.45-50 Ultraviolet A (UVA) ljóseindir oxandi viðbrögð með því að örva innra ljósnæmisvaka, svo sem porfyrín, NADPH oxidasa og ríbóflavín. 8-Oxo-7,8-díhýdródúanín (8-oxoGua) er aðal UVA-miðlað DNA oxunarafurðin sem myndast við oxun OH-röntgen, 1-rafeindarefnisoxíð og einfalt súrefni sem aðallega bregst við guanín. 51 Myndun guaníns róttækrar katjónar í einangruðri DNA hefur verið sýnt fram á að það komi fram með beinum áhrifum jónandi geislunar. 52,53 Eftir útsetningu fyrir jónandi geislun minnkar innanfrumuþéttni glútaþíon (GSH) til skamms tíma en eykst síðan aftur.54
Þungmálmar jónir
Þungur málmjónir, eins og járn, kopar, kadmíum, kvikasilfur, nikkel, blý og arsen, geta valdið myndun hvarfefna og veldur frumuskemmdum með því að eyða ensímvirkni með peroxíðmyndun og viðbrögð við kjarnapróteinum og DNA.55
Eitt af mikilvægustu aðferðum málmleiddra kynhneigðra kynslóða er með Fenton-gerð viðbrögðum. Superoxíð jón og vetnisperoxíð geta haft samskipti við umskipti málma, svo sem járn og kopar, í gegnum málm hvataða Haber-Weiss / Fenton viðbrögð við myndun OH róttamanna.
Metal31 1 $ O2 / Metal21 1 O2 Haber Weiss Metal21 1 H2 O2 / Metal31 1 OH 2 1 $ OH Fentónviðbrögð
Fyrir utan Fenton-gerð og Haber-Weiss-gerð kerfi geta ákveðnar málmjónir bregst beint við frumu sameindir til að mynda sindurefnahvarf, svo sem þíóólstakefni, eða örva frumuskiptingarleiðir. Þessir róttækar geta einnig brugðist við öðrum þíólólmoleknum til að mynda O22 .. O22. er breytt í H2O2, sem veldur viðbótar súrefni róttækri kynslóð. Sumir málmar, eins og arsenít, örva ROS myndun óbeint með virkjun á raðaframleiðslukerfum í frumum. 56
Arsen er mjög eitrað þáttur sem framleiðir margs konar ROS, þar með talin súrefni (O2 2), singlet súrefni (1O2), peroxýlstakeindir (ROO), köfnunarefnisoxíð (NO), vetnisperoxíð (H2O2) og dímetýlsínínperoxýlstakar CH3) 2AsOO] .57-59 Arsen (III) efnasambönd geta hamlað andoxunarefnum ensímum, sérstaklega GSH-háðum ensímum, svo sem glútaþíon-S-transferasa (GST), glútaþíon peroxidasa (GSH-Px) og GSH-redúktasa, með bindiefni - Súlfhýdrýl (-SH) hópar þeirra. 60,61
Leiða eykur fituprótíðun. 62 Veruleg lækkun á starfsemi vefja SOD og heila GPx hefur verið tilkynnt eftir blýi útsetningu. 63,64 Skipti á sinki, sem virkar sem samvirkni í mörgum ensímum með blýi, leiðir til þess að slökkva á slíkum ensímum. Leiðaáhrif geta valdið hömlun á GST með því að hafa áhrif á vefjarþíól.
ROS sem myndast af málmkatalystum viðbrögðum getur mótað DNA basa. Þrjár grunnarskiptingar, G / C, G / T og C / T, geta komið fram vegna oxunar skemmda af málmjónum, svo sem Fe21, Cu21 og Ni21. Reid et al65 sýndi að G / C var aðallega framleitt með Fe21 meðan C / T skipting var með Cu21 og Ni21.
ANTIOXIDANTS
Mannslíkaminn er búinn með fjölbreyttu andoxunarefnum sem stuðla að því að móta áhrif oxandi efna. Fyrir alla hagnýta tilgangi er hægt að skipta þessum efnum í 2 flokkana: ensímatækni (tafla 2) og nonenzymatic (Tafla 3).



Enzymatic andoxunarefni
Helstu ensímandi andoxunarefnin í lungum eru SOD (EC 1.15.1.11), katalasa (EC 1.11.1.6) og GSH-Px (EC 1.11.1.9). Í viðbót við þessar helstu ensím, önnur andoxunarefni, þar á meðal Heme oxygenasi-1 (EC 1.14.99.3), og redox prótín, eins og thioredoxins (TRXs, EC 1.8.4.10), peroxiredoxins (PRXs, EC 1.11.1.15), og glutaredoxins, hafa einnig verið fundið að gegna mikilvægu hlutverki í lungum andoxunarefni varnir.
Þar sem superoxíð er aðal ROS framleitt úr ýmsum aðilum er dismutation hennar með SOD aðallega mikilvæg fyrir hverja frumu. Öll 3 form SOD, það er, CuZn-SOD, Mn-SOD og EC-SOD, eru víða tjáð í lungum manna. Mn-SOD er staðbundin í hvatbera matrinu. EC-SOD er fyrst og fremst staðbundin í utanfrumuformi, sérstaklega á svæðum sem innihalda mikið magn af tegund I kollagenfrumum og kringum lungna- og kerfisbundna skip. Það hefur einnig fundist í berkjuþekju, alveolar epithelium og alveolar macrophages.66,67 Á heildina litið er CuZn-SOD og Mn-SOD yfirleitt talið virka sem magnhreinsiefni superoxíðrauka. Tiltölulega hátt EC-SOD stig í lungum með sértæka bindingu við utanfrumuvökvaþættirnar geta verið grundvallarþáttur lungnagreininga. 68
H2O2, sem er framleitt með virkni SOS eða virkni oxýasa, eins og xantínoxíð, er dregið úr vatni með hvata og GSH-Px. Catalase er til staðar sem tetra- mer samanstendur af 4 sams konar einliða, sem hver um sig inniheldur heilahóp á virkum stað. Niðurbrot H2O2 er náð með umbreytingu á 2 byggingu katalasa-ferricatalasa (járn samræmd í vatni) og samsetning I (járn flókið með súrefnisatóm). Catalase binst einnig NADPH sem minnkandi jafngildi til að koma í veg fyrir oxunarvirkjun ensímsins (myndun efnasambands II) með H2O2 eins og það er dregið úr í vatn. 69
Ensím í redox hringrás sem ber ábyrgð á því að draga úr H2O2 og lípíða í hydroperoxides (mynda sem afleiðing af himnu lípíð peroxíderingu) fela í sér GSH-Pxs.70 The GSH-Pxs eru á fjöldamörgum tetrameric ensím sem innihalda einstaka amínósýru selenocysteine innan virkum stöðum og nota þiól með lág mólmassa, svo sem GSH, til að draga úr H2O2 og fituefnum peroxíðum við samsvarandi alkóhól. GSH-Px-1 (cellular GSH-Px) er fjallað um fjórar GSH-Px, og dregur úr H2O2 og fitusýruperoxíðum, en ekki esteruð peroxýl lipíð. 71 Esterified fituefni eru minnkuð með himnubundnu GSH -Px-4 (fosfólípíðhýdroxýoxíð GSH-Px), sem hægt er að nota nokkrar mismunandi thiól með lág sameindaþyngd sem að minnka jafngildi. GSH-PX-2 (meltingarvegi GSH-Px) er staðbundin í meltingarvegi þekjufrumum þar sem það þjónar að draga úr mataræði peroxides.72 GSH-PX-3 (utanfrumuvökva GSH-PX) er eini meðlimur GSH-PX fjölskyldu sem býr í utanfrumuhólfinu og er talið vera eitt mikilvægasta utanfrumu andoxunarefnið í spendýrum. Af þeim er rannsakað utanfrumugerð GSH-Px í mönnum lungum. 73
Að auki tengist förgun H2O2 náið tengslum við nokkrar þíól-innihaldandi ensím, þ.e. TRXs (TRX1 og TRX2), tioredoxín-redúktasa (EC 1.8.1.9) (TRRs), PRXs (sem eru tioredoxinperoxidasi) og glútaroxín. 74
Tvær TRX og TRR hafa verið einkennist í frumum úr mönnum, sem eru fyrir hendi bæði í cýtosól og hvatberum. Í lungum eru TRX og TRR tjáð í berkju- og alveolarþekju og makrólfur. Sex mismunandi PRXs hafa fundist í mönnum frumum, mismunandi í öfgafræðilegum hólfum þeirra. Tilraunagreinarannsóknir hafa leitt í ljós mikilvægi PRX VI við verndun alveolar epithelium. Human lungum tjáir allar PRXs í berkjuþekju, alveolar þekjuvef, og macrophages.75 PRX V hefur nýlega fundist til að virka eins og a peroxýnítrít afoxunarmiðlinum, 76 sem þýðir að það geta virkað sem hugsanleg verndandi efnasambandi í þróun ROS-miðlað lungnameiðslum .77
Algengt fyrir þessar andoxunarefni er krafa NADPH sem minnkandi jafngildi. NADPH heldur catalase í virka forminu og er notað sem cofactor með TRX og GSH redúktasa (EC 1.6.4.2), sem umbreytir GSSG til GSH, samhliða undirlagi fyrir GSH-Px. Innrennsli NADPH er síðan myndað af lækkun NADP1 með glúkósa-6-fosfat dehýdrógenasa, fyrsta og takmörkunarsýru ensímsins í pennu-fosfatleiðinni, við umbreytingu glúkósa-6-fosfats í 6-fosfóglúkónólaktón. Með því að mynda NADPH er glúkósa-6-fosfat dehýdrógenasi mikilvægur þáttur í cýtósósól-GSH-stuðningsgetu (GSH / GSSG) og því er hægt að líta svo á að það sé nauðsynlegt, reglulegt andoxunarefni ensím. 78,79
GST (EC 2.5.1.18), annar andoxunarefni ensím fjölskylda, gera óvirkan efnaskipta, eins og ómettað aldehýð, epoxíð og vatnsperoxíð. Þrjár helstu fjölskyldur GSTs hefur verið lýst: umfrymi GST, hvatberum GST, 80,81 og himnutengds netbólu GST sem hefur hlutverki í eicosanoid og GSH metabolism.82 Sjö flokkar umfrymi GST eru tilgreindir í spendýrum, tilnefnd Alpha, Mu, Pi, Sigma, Theta, Omega og Zeta.83-86. Meðan á óþvinguðum kringumstæðum stendur, skiptast á flokkur Mu og Pi GSTs við kínasa Ask1 og JNK, í sömu röð, og hindra þessar kínasa. 87-89. Sýnt hefur verið fram á að GSTP1 dissociates frá JNK í svörun við oxandi streitu. 89 GSTP1 hefur einnig líkamlega áhrif á PRX VI og leiðir til endurheimtar PRX ensímvirkni með glutathionýlering á oxuðu próteinum. 90
Nonenzymatic andoxunarefni
Nonenzymatic andoxunarefnum innihalda lítinn sameindarþyngd efnasambönd, eins og vítamín (vítamín C og E), b-karótín, þvagsýru og GSH, þrígeptíð (Lg-glútamýl-L-sýsteinýl-L-glýsín) sem samanstanda af þíól súlfhýdrýl) hópur.
C-vítamín (askorbínsýra)
Vatnsleysanlegt C-vítamín (askorbínsýra) veitir andoxunareiginleika vatnsfasa í frumum og utanfrumum, einkum með því að hreinsa súrefnisstorkuefni. Það breytir vítamínum E vítamínum til E-vítamíns. Plasmaþéttni þess hefur reynst lækka með age.91,92
E-vítamín (a-tókopólól)
Lipid-leysanlegt E-vítamín er þétt á vatnsfælna innri vef frumuhimnu og er aðalvarnarefni gegn skaða á oxunarefnum. E-vítamín gefur rafeind til peroxýlstakeindar, sem er framleitt meðan á peroxíðun stendur. a-Tókóferól er mest virka form E-vítamíns og helstu himnutengdu andoxunarefni í frumu. E-vítamín veldur apoptosis á krabbameinsfrumum og hindrar myndun rétthyrninga. 93
Glútaþíon
GSH er mjög mikið í öllum klefahólfum og er aðal leysanlegt andoxunarefni. GSH / GSSG hlutfallið er mikilvægur þáttur í oxunarálagi. GSH sýnir andoxunarefni þess á nokkra vegu. 94 Það detoxifies vetnisperoxíð og lípíðperoxíð með aðgerð GSH-Px. GSH leggur rafeindið í H2O2 til að minnka það í H2O og O2. GSSG er aftur minnkað í GSH með GSH redúktasa sem notar NAD (P) H sem rafeind gjafa. GSH-Px eru einnig mikilvæg til að vernda frumuhimnu úr peroxíðmyndun. Minni glútaþíon veitir róteindir til himnafitu og verndar þau gegn oxunarárásum. 95
GSH er cofactor fyrir nokkrar afeitandi ensím, svo sem GSH-Px og transferasa. Það hefur hlutverk í að umbreyta C-vítamín og E til baka í virku formi þeirra. GSH verndar frumum gegn apoptosis með samskiptum við proapoptotic og antiapoptotic skilunarleiðir. 94 Það stjórnar og virkjar einnig nokkrar uppskriftarþættir, svo sem AP-1, NF-kB og Sp-1.
Karótenóíðum (b-karótín)
Karótenóíur eru litarefni sem finnast í plöntum. Fyrst og fremst hefur b-karótín reynst að hvarfast við peroxýl (ROO), hýdroxýl (OH) og superoxíð (O22.) Radíus. 96 Karótenóíð sýna andoxunaráhrif sín í lágþrýstingsþrýstingi en geta haft foroxunaráhrif við hærri súrefni styrkur. 97 Bæði karótenóíð og retínósýrur (RAs) geta stjórnað umritunarmörkum. 98 b-karótín hamlar oxunareflandi NF-kB virkjun og interleukin (IL) -6 og æxlisfrumaþáttur-framleiðslu. Karótenóíur hafa einnig áhrif á frumufjölgun frumna. Víxlverkunaráhrif RA hafa verið sýndar í nokkrum rannsóknum. Þessi áhrif af RA er miðlað aðallega af retínósýruviðtökum og eru mismunandi eftir tegundum frumna. Í brjóstkrabbameinsfrumum var sýnt fram á retínósýruviðtaka að valda vaxtarhömlun með því að örva handtöku frumna, apoptosis eða bæði. 99,100
ÁHRIF SJÁLFREYNDAR STRESS: Erfðafræðileg, lífeðlisfræðileg og lífefnafræðileg vélbúnaður
Oxunartruflanir eiga sér stað þegar jafnvægi milli andoxunarefna og ROS er truflað vegna þess að það er annaðhvort niðurbrot af andoxunarefnum eða uppsöfnun ROS. Þegar oxandi álag kemur fram, reyna frumur að vinna gegn oxunarefnum og endurheimta rauðkaljubalann með því að virkja eða slökkva á genum sem kóða varnar ensím, transcription factors og byggingarprótein. 101,102 Hlutfall milli oxaðs og minnkaðs glútaþíon (2GSH / GSSG) er eitt af mikilvægum þáttum oxandi streitu í líkamanum. Hærri framleiðsla á ROS í líkamanum getur breytt DNA-uppbyggingu, leitt til breytinga á próteinum og fitum, virkjun nokkurra þunglyndisþáttaþátta og framleiðslu á bólgueyðandi og bólgueyðandi frumudrepum.
Áhrif oxandi streitu á DNA
ROS getur leitt til breytinga á DNA á nokkra vegu, sem felur í sér niðurbrot basa, ein- eða tvístrengdar DNA brot, púrín, pýrimídín eða sykurbundnar breytingar, stökkbreytingar, eyðingar eða translocations og víxlbindingu við prótein. Flest þessara DNA breytinga (mynd 1) eru mjög viðkvæm fyrir krabbameinsvaldandi áhrifum, öldrun og taugakvilla, hjarta- og æðasjúkdóma og sjálfsónæmissjúkdóma. Tóbak reyk, redox málmar og nonredox málmar, svo sem járn, kadmíum, króm og arsen, taka einnig þátt í krabbameinsvaldandi áhrifum og öldrun með því að mynda sindurefna eða bindast með þíól hópum. Myndun 8-OH-G er þekktasti DNA skaði sem kemur fram með oxunarálagi og er hugsanlegt lífvera fyrir krabbameinsvaldandi áhrif.

Promoter svæðum gena innihalda samstöðu röð fyrir umritun þætti. Þessar umritunarþættir bindandi staður innihalda GC-ríkur röð sem eru næmir fyrir oxun árásum. Myndun 8-OH-G DNA á bindisþáttum bindingarþáttar getur breytt bindingu uppritunarþátta og þannig breytt tjáningu tengdra gena eins og sýnt hefur verið fyrir AP-1 og Sp-1 miðunarmörk. 103 Að auki 8-OH-G, 8,59-sýkló-29-deoxýadenósín (sýkló-dA) hefur einnig verið sýnt fram á að hindra uppskrift frá blaðamaður geni í frumukerfi ef hún er staðsett í TATA-kassa. 104 TATA-bindandi próteinin hefst áritun með því að breyta beygingu DNA. Binding TATA-bindandi próteins getur verið skert vegna nærveru sýkló-dA.
Oxandi streitu veldur óstöðugleika örsatellíts (stutt tíðni endurtekningar) svæði. Redox virk málmjónir, hýdroxýlstakeindir auka óstöðugleika í óstöðugleika. 105 Þrátt fyrir að einangruð DNA brot, sem orsakast af oxunarefnum, geta auðveldlega þolist af frumum, geta tvöfaltstrengar DNA brot, sem orsakast af jónandi geislun, verið veruleg ógn fyrir lifun lifrarinnar. 106
Metýlering á CpG-eyjum í DNA er mikilvægur epigenetísk vélbúnaður sem getur leitt til genþols. Oxun 5-MeCyt við 5-hýdroxýmetýlúracil (5-OHMeUra) getur átt sér stað með afmengun / oxunarviðbrögðum af týmíni eða 5-hýdroxýmetýlcytósín milliefni. 107 Auk þess að mótmæla genþrýstingin virðist DNA metýlering einnig hafa áhrif á chromatín organization.108 Aberrant DNA methylation mynstur framkölluð af oxunarárásum hafa einnig áhrif á DNA viðgerð virkni.
Áhrif oxandi streitu á fitu
ROS getur valdið lipid peroxidation og truflað frumuhimnubúnaðinn sem getur hindrað himnutengda viðtaka og ensíma og aukið vefnæmi. 109 Vörur af fituefnumoxun, eins og MDA og ómettað aldehýð, geta gert margar frumuprótín óvirkar með því að mynda próteinhvert tengsl.110-112 4-Hýdroxý-2-nonenal veldur niðurbroti innanfrumu GSH og hvetur til peroxíðsframleiðslu, 113,114 virkjar húðþekjuviðtakaviðtaka, 115 og veldur framleiðslu fibrónektíns. 116 Lipid peroxidation vörur, svo sem ísópróstanar og þíóbíkbitúrsýru hvarfefni , hafa verið notuð sem óbein lífmerki af oxunarálagi og aukin gildi voru sýnd í útöndunarsamdrætti eða berkjuveiruvökvavökva eða lungum sjúklinga með langvarandi lungnateppu eða reykingamenn .117-119
Áhrif oxandi streitu á próteinum
ROS getur valdið sundrungu á peptíðkeðjunni, breytingu á rafhleðslu próteina, víxlverkun próteina og oxun tiltekinna amínósýra og leitt því til aukinnar næmni fyrir próteinlosun með niðurbroti með sérstökum próteasahemlum. 120 Cysteín og metíónínleifar í próteinum eru sérstaklega næmari fyrir oxidation.121 fyrir oxun á súlfhýdrýl hópa eða meþiónfnleifar af próteinum valdið stellingabreytingar, prótein þróast, og degradation.8,121-123 ensím sem hafa málma á eða nálægt virku vefsvæði þeirra eru sérstaklega næmari fyrir málm catalyzed oxun. Sýnt hefur verið fram á að oxandi áhrif ensíma hafa áhrif á starfsemi þeirra. 124,125
Í sumum tilfellum getur sérstakt oxun próteina átt sér stað. Til dæmis má metíónín oxa metíónínsúlfoxíð126 og fenýlalanín til o-týrósín127; Súlfhýdrýlhópar geta verið oxaðir til að mynda tvísúlfíðbindingar; 128 og karbónýlhópar má kynna í hliðarkeðjur próteina. Gamma geislar, málmkatalysandi oxun, HOCl og óson geta valdið myndun karbónýlhópa.129
Áhrif oxandi streitu á segamyndun
ROS getur valdið tjáningu nokkurra gena sem taka þátt í segulmyndun. 1,130 Hátt hlutfall GSH / GSSG er mikilvægt til að vernda frumuna gegn oxunarefnum. Slökun á þessu hlutfalli veldur virkjun rauðkornaviðkvæmra umritunarþátta, svo sem NF-kB, AP-1, kjarnorkuþáttur virkjunar T-frumna og blóðoxíð-örvandi þáttur 1, sem taka þátt í bólguviðbrögðum. Virkjun áritunarþátta í gegnum ROS er náð með því að nota merkjatransduction cascades sem senda upplýsingarnar utan frá til frumu. Týrósín kínasaviðtaka, flestir vaxtarþáttarviðtaka, eins og vaxtarþáttur viðtaka í húðþekju, vaxtarþáttarviðtaka í æðar og viðtaka fyrir vaxtarþáttur blóðflagna, prótein týrósín fosfatasa og serín / þreónín kínasa eru markmið ROS.131-133 Auka-frumuveirukerfisbundnar kínasa, JNK og p38, sem eru meðlimir mítógenvirkja prótína kínasa fjölskyldu og taka þátt í nokkrum ferlum í frumu, þar með talið fjölgun, frávik og apoptosis, geta einnig verið stjórnað af oxunarefnum.
Undir oxunarástandi eru cysteinleifar í DNA-bindingarstöðum c-júní, sumar AP-1 undireiningar og hindrandi kB kínasi gegnt reversible S-glutathiolation. Glutaredoxin og TRX hafa verið tilkynnt að gegna mikilvægu hlutverki í reglum um rauðkornaviðkvæm merki, eins og NF-kB og AP-1, p38 mítógenvirkja prótínkínasa og JNK.134-137
NF-kB getur verið virkjað til að bregðast við oxun streitu skilyrði, svo sem ROS, sindurefna og UV geislun. 138 fosfórun IB losnar NF-kB og gerir það kleift að komast inn í kjarnann til að virkja genskiptingu. 139 Greint hefur verið frá fjölda kinasa til að fosfórýlera IkBs við serínleifarnar. Þessar kínasa eru markmið oxunarmerkja til að virkja NF-kB.140 Reduce Agents auka NF-kB DNA bindingu, en oxandi efni hamla DNA bindingu NF-kB. TRX getur beitt 2 gagnstæðum aðgerðum í reglugerð NF-kB: í frumumæxlinu blokkar það niðurbrot á IkB og hamlar NF-kB virkjun en eykur NF-kB DNA bindingu í kjarnanum. 141 Virkjun NF-kB gegnum oxunar tengd niðurbrot af IkB leiðir til virkjunar nokkurra varnarefna sem tengjast andoxunarefni. NF-kB stjórnar tjáningu nokkurra gena sem taka þátt í ónæmissvörun, svo sem IL-1b, IL-6, æxlisfrumnaþáttur-a, IL-8 og nokkrar viðloðunarsameindir. 142,143 NF-kB stjórnar einnig æðamyndun og fjölgun og aðgreining frumna.
AP-1 er einnig stjórnað með redox ástandi. Í nærveru H2O2 geta sum málmjónir örvað virkjun AP-1. Aukning á hlutfalli GSH / GSSG eykur AP-1 bindingu meðan GSSG hamlar DNA-bindingu AP-1.144 DNA bindingu Fos / Jun heteródímerans er aukin með því að draga úr einum varðveitt cysteine í DNA bindandi léni hvers próteinin, 145 en DNA-bindingu AP-1 má hamla með GSSG í mörgum frumum, sem bendir til þess að myndun díúlfíðbindinga með cysteínleifar hamlar AP-1 DNA bindingu. 146,147 Merkjatransduction via oxidative stress is summarized in Figure 2.

Ályktanir
Oxandi streita getur stafað af offramleiðslu ROS með efnaskiptaviðbrögðum sem nota súrefni og breyta jafnvægi á milli oxandi /andoxunarefni Staða í þágu oxunarefna. ROS eru framleidd með frumueyðandi starfsemi og umhverfisþáttum, svo sem loftmengunarefni eða sígarettureykur. ROS eru mjög hvarfgjarn sameindir vegna ópaðar rafeinda í uppbyggingu þeirra og bregðast við nokkrum líffræðilegum macromolecules í klefi, eins og kolvetni, kjarnsýrur, fituefni og prótein og breyta virkni þeirra. ROS hefur einnig áhrif á tjáningu nokkurra gena með því að breyta reglum um redox-viðkvæma uppskrift og endurmyndun litabreytinga með breytingu á histón acetýlering / deacetylering. Reglugerð um redox er mikilvægt fyrir lífvænleika, virkjun, fjölgun og líffæravirkni.
Tilvísanir:
HEIMILDIR
1. Valko M, Rhódos CJ, Moncol J, Izakovic M, Mazur M. Frídreifar, málmar og andoxunarefni í krabbameini með oxunarálag. Chem Biol Interact. 2006; 160: 1-40.
2. Halliwell B, Gutteridge JMC. Free radicals í líffræði og læknisfræði. 3rd ed. New York: Oxford University Press; 1999.
3. Marnett LJ. Lipid peroxidationdDNA skemmdir af malondialdehýð. Mutat Res. 1999; 424: 83-95.
4. Siems WG, Grune T, Esterbauer H. 4-Hydroxynonenal myndun við blóðþurrð og endurblöndun smáþarma í rottum. Life Sci. 1995; 57: 785–789.
5. Stadtman ER. Hlutverk oxandi tegunda við öldrun. Curr Med Chem. 2004; 11: 1105-1112.
6. Wang MY, Dhingra K, Hittelman WN, Liehr JG, deAndrade M, Li DH. Lipid peroxidation-framkölluð hugsanleg malondialdehýð-DNA adducts í brjóstvefi manna. Krabbamein Epidemiol Biomarkers Prev. 1996; 5: 705-710.
7. Jenner P. Oxidative streita í Parkinsonsveiki. Ann Neurol. 2003; 53: S26-S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Mat á oxunarskemmdum á próteinum, fitum og DNA í heilanum frá sjúklingum með Alzheimerssjúkdóm. J Neurochem. 1997; 68: 2061-2069.
9. Sayre LM, Smith MA, Perry G. Efnafræði og lífefnafræði oxunar streitu í taugahrörnunarsjúkdómum. Curr Med Chem. 2001; 8: 721-738.
10. Toshniwal PK, Zarling EJ. Vísbendingar um aukin fituprepun í mænusigg. Neurochem Res. 1992; 17: 205-207.
11. Dhalla NS, Temsah RM, Netticadan T. Hlutverk oxandi streitu í hjarta- og æðasjúkdómum. J Hypertens. 2000; 18: 655-673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, et al. Rannsókn á oxunarálagi í rottum fyrir langvarandi heilahimnuspennu. Neurochem Int. 2005; 46: 601-611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Framleiðsla superoxíðanjóns er aukin í líkani af erfðaháþrýstingi: hlutverk endothelsins. Háþrýstingur. 1999; 33: 1353-1358.
14. Kukreja RC, Hess ML. Súrefnisfrjálst róttæka kerfið: frá jöfnum í gegnum himna-prótein milliverkanir við hjarta- og æðasjúkdóma og vernd. Cardiovasc Res. 1992; 26: 641-655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, et al. Reykingar á sígarettu stuðla að aukningu á oxandi DNA skaða, 8-hýdroxýdeoxýgúanósíni, á miðlægum stað lungnasjúkdómsins. Krabbameinsvaldandi áhrif. 1997; 18: 1763-1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Oxandi og nítrósandi atburðir í astma. Free Radic Biol Med. 2003; 35: 213-225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, et al. Fylgni á skömmtum superoxid dismutasa skorts á loftflæði hindrun í astma. Am J Respir Crit Care Med. 2005; 172: 306-313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, et al. Óvirkjun á oxíðdíoxýdesmúdasa í sjúkdómsgreiningu astmahugsunar og endurvirkni. Er J Pathol. 2005; 166: 663-674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Oxidative streita og þættir hennar í öndunarvegi barna með astma. Ofnæmi. 2008; 63: 1605-1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, et al. Oxandi streitu og erfða- og faraldsfræðilegir ákvarðanir á oxunarefnum í astma astma. J Allergy Clin Immunol. 2006; 118: 1097-1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Alvarlegt astma rannsóknaráætlun. Hjartastöðugleiki glútaþíon er breytt hjá börnum með alvarlega astma: vísbendingar um oxunartruflanir. J Allergy Clin Immunol. 2009; 123: 146-152.
22. Miller DM, Buettner GR, Aust SD. Flutningur málma sem hvatar við "sjálfstætt oxun" viðbrögð. Free Radic Biol Med. 1990; 8: 95-108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, Dème D, Pommier J. Mechanism af vetnisperoxíðmyndun hvattur af NADPH oxidasa í skjaldkirtilsplasma himnu. J Biol Chem. 1991; 266: 3739-3743.
24. Granger DN. Hlutverk xanthínoxíðasa og kyrningafjölskylda í meiðslum vegna meinvarpsfrumna. Am J Physiol. 1988; 255: H1269-H1275.
25. Fenton HJH. Oxun vínsýru í viðurvist járns. J Chem Soc. 1984; 65: 899-910.
26. Haber F, Weiss JJ. Katalískar niðurbrot vetnisperoxíðs með járnsöltum. Proc R Soc Lond Ser A. 1934; 147: 332-351.
27. Liochev SI, Fridovich I. The Haber-Weiss cycled70 árum síðar: valmynd. Redox Rep. 2002; 7: 55-57.
28. Klebanoff SJ. Myeloperoxidasi: vinur og fjandmaður. J Leukoc Biol. 2005; 77: 598-625.
29. Whiteman M, Jenner A, Halliwell B. Hypochlorous sýruvaldandi grunnbreytur í einangruðum kálfþímus DNA. Chem Res Toxicol. 1997; 10: 1240-1246.
30. Kulcharyk PA, Heinecke JW. Hýpróklórsýra, framleitt af myeloperoxidasa kerfinu af fagfrumum úr mönnum, veldur samgildum víxlbindingum milli DNA og próteins. Lífefnafræði. 2001; 40: 3648-3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Song W, et al. Sagan af tveimur deilum: skilgreinir bæði hlutverk peroxidasa við nitrotyrosín myndun in vivo með því að nota eosinophil peroxidasa og myeloperoxidasdeficient mýs og eðli peroxidasa mynda viðbrögð köfnunarefnis tegundir. J Biol Chem. 2002; 277: 17415-17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, et al. Mjög mikil eosinophil degranulation og peroxidasa miðluð oxun í öndunarvegi prótein koma ekki fram í mótefnavökva með músum ovalbumin-áskorun lungnabólgu. J Immunol. 2001; 167: 1672-1682.
33. Van Dalen CJ, Winterbourn CC, Senthilmohan R, Ketill AJ. Nitríti sem hvarfefni og hemill myeloperoxidasa. Áhrif á nitrunar- og hýdróklórsýruframleiðslu á bólusvæðum. J Biol Chem. 2000; 275: 11638-11644.
34. Wood LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Lipid peroxidation, sem ákvarðað er með plasma ísópróstani, tengist alvarleika sjúkdóms við væga astma. Fituefni. 2000; 35: 967-974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Aukin 8-ísópróstan, merki um oxunarálag, í útöndunarsamdrætti astma sjúklinga. Am J Respir Crit Care Med. 1999; 160: 216-220.
36. Kirkja DF, Pryor WA. Frí-róttæk efnafræði sígarettureykur og eiturefnafræðileg áhrif þess. Environ Health Perspect. 1985; 64: 111-126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, et al. Ósjálfstætt bólga metið í sputum og berkjuvökvavökva frá astmatískum lyfjum: nýtt noninvasive tól í faraldsfræðilegum rannsóknum á loftmengun og astma. Free Radic Biol Med. 1999; 27: 1448-1454.
38. Nightingale JA, Rogers DF, Barnes PJ. Áhrif innöndunar ósons við útöndun nituroxíðs, lungnastarfsemi og framkallað sputum hjá eðlilegum og astmískum einstaklingum. Þorax. 1999; 54: 1061-1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, et al. Redox virkni loftfrumna agna á mismunandi stöðum í Los Angeles Basin. Environ Res. 2005; 99: 40-47.
40. Comhair SA, Thomassen MJ, Erzurum SC. Mismunandi framköllun á utanfrumu glútaþíon peroxidasa og nituroxíðsýnatasa 2 í öndunarvegi hjá heilbrigðum einstaklingum sem verða fyrir 100% O (2) eða sígarettureyk. Am J Respir Cell Mol Biol. 2000; 23: 350-354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Oxidant-miðlað lungnaskemmdir í bráðri öndunarerfiðleikum. Crit Care Med. 1999; 27: 2028-2030.
42. Biaglow JE, Mitchell JB, Held K. Mikilvægi peroxíðs og superoxíðs í röntgenviðbrögðum. Int J Radiat Oncol Biol Phys. 1992; 22: 665-669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Koparjónmótað næmi á kjarnaklefa tengibúnaði við jónandi geislun. Lífefnafræði. 1993; 32: 6214-6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Alfa agnir hefja líffræðilega framleiðslu á superoxíð anjón og vetnisperoxíði í frumum manna. Krabbamein Res. 1997; 57: 3963-3971.
45. Tuttle SW, varar ME, Mitchell JB, Biaglow JE. Næmi fyrir efnafræðilegum oxunarefnum og geislun í CHO frumu línum sem eru ófullnægjandi í oxandi pentósa hringrás starfsemi. Int J Radiat Oncol Biol Phys. 1992; 22: 671-675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, et al. Mangan
superoxid dismutasa-miðlað gen tjáningu í geislunarvirkni
aðlögunarhæfar viðbrögð. Mol Cell Biol. 2003; 23: 2362-2378.
47. Azzam EI, de Toledo SM, Spitz DR, Little JB. Oxandi efnaskipti
modulates merki transduction og micronucleus myndun í andstæðingi
frumur úr geislameðhöndluðum eðlilegum trefjum úr mönnum. Krabbamein Res.
2002; 62: 5436-5442.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Ionizing geislun framkölluð, hvatbera háð kynslóð af viðbrögðum
súrefni / köfnunarefni. Krabbamein Res. 2001; 61: 3894-3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK leið í
geislunarviðbrögð. Oncogene. 2003; 22: 5885-5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, et al. Thioredoxin
kjarnaútfelling og samskipti við redox-þáttur-1 virkjar AP-1 uppskriftarþáttinn sem svar við jónandi geislun. Krabbamein Res. 2000; 60: 6688-6695.
51. Cadet J, Douki T, Gasparutto D, Ravanat JL. Oxandi skemmdir á DNA: myndun, mælingar og lífefnafræðilegir eiginleikar. Mutat Res. 2003; 531: 5-23.
52. Yokoya A, Cunniffe SM, O'Neill P. Áhrif vökva á virkjun á brotum á strandar og grunnskemmdir í plasmíð DNA kvikmyndum með gammaradiation. J er Chem Soc. 2002; 124: 8859-8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Viðbrögð frumna og vefja við oxunarskaða. Lab Invest. 1993; 69: 261-274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, et al. Kjarnaþáttur kappa B í kjölfarið er háð framköllun gamma glutamýlcysteine synthetasa með jónandi geislun í T98G manna glioblastoma frumum. Free Radic Biol Med. 1998; 24: 1256-1268.
55. Stohs SJ, Bagchi D. Oxunarefni í eiturhrifum málmjóna. Free Radic Biol Med. 1995; 18: 321-336.
56. Leonard SS, Harris GK, Shi X. Metal-framkallað oxandi streitu og merki ummyndun. Free Radic Biol Med. 2004; 37: 1921-1942.
57. Shi H, Shi X, Liu KJ. Oxandi verkun eiturhrifa arsen og krabbameinsvaldandi áhrif. Mol Cell Biochem. 2004; 255: 67-78.
58. Pi J, Horiguchi S, Sól Y, Nikaido M, Shimojo N, Hayashi T. Möguleg aðferð við skerðingu á myndun nituroxíðs vegna langvarandi útsetningar fyrir arsenati í kanínum. Free Radic Biol Med.2003; 35: 102-113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. DNAstrand hlé sem valdið er af dímetýlsínsýru, umbrotsefni ólífrænna arsenafræðilegra efna, er sterklega aukið með radoxíði afoxíðs. Biol Pharm Bull. 1995; 18: 45-58.
60. Waalkes MP, Liu J, Ward JM, Diwan LA. Kerfi undirliggjandi arsen krabbameinsvaldandi: Ofnæmi fyrir músum sem verða fyrir ólífrænum arseni meðan á meðgöngu stendur. Eiturefnafræði. 2004; 198: 31-38.
61. Schiller CM, Fowler BA, Woods JS. Áhrif arsens á virkni pyruvat dehýdrógenasa. Environ Health Perspect. 1977; 19: 205-207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Inntaka frjálsra raðra í taugafrumum og leiðbólgu. Mol Cell Biochem. 1991; 103: 73-83.
63. Tripathi RM, Raghunath R, Mahapatra S. Blóðleiða og áhrif þess á Cd, Cu, Zn, Fe og blóðrauðagildi barna. Sci Total Environ. 2001; 277: 161-168.
64. Nehru B, Dua R. Áhrif mataræði selen á eiturverkunum á taugakerfi. J Environ Pathol Toxicol Oncol. 1997; 16: 47-50.
65. Reid TM, Feig DI, Loeb LA. Stökkbreytingar af málmum völdum súrefnisstorkum. Environ Health Perspect. 1994; 102 (suppl 3): 57-61.
66. Kinnula VL, Crapo JD. Superoxíðdísútamasa í lungum og lungnasjúkdómum í mönnum. Am J Respir Crit Care Med. 2003; 167: 1600-1619.
67. Kinnula VL. Framleiðsla og niðurbrot súrefnis umbrotsefna í bólgusjúkdómi í lungum í mönnum. Curr Drug Markmið Inflamm Ofnæmi. 2005; 4: 465-470.
68. Zelko IN, Mariani TJ, Folz RJ. Superoxíð tvímóteasa fjölgene fjölskylda: samanburður á CuZn-SOD (SOD1), Mn-SOD (SOD2) og EC-SOD (SOD3) genarefnum, þróun og tjáningu. Free Radic Biol Med. 2002; 33: 337-349.
69. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Aðferðir við verndun katalasa af NADPH. Kinetics og stoichiometry. J Biol Chem. 1999; 274: 13908-13914.
70. Flohé L. Glutathione peroxidasa. Basic Life Sci. 1988; 49: 663-668.
71. Arthur JR. The glútaþíon peroxíðasa. Cell Mol Life Sci. 2000; 57: 1825-1835.
72. Chu FF, Doroshow JH, Esworthy RS. Tjáning, einkenni og vefja dreifingu nýrra sellulíns háð glútaþíon peroxidasa, GSHPx-GI. J Biol Chem. 1993; 268: 2571-2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Örkyrningafæð af glútaþíon peroxidasa í astma lungum: vísbendingar um rauðkornaþéttni tjáningar í frumum í frumum í mönnum. FASEB J. 2001; 15: 70-78.
74. Gromer S, Urig S, Becker K. Thioredoxin kerfi frá vísindum til heilsugæslustöðvar. Með endurb. 2004; 24: 40-89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, Pääkkö P, o.fl. Frumgreinandi tjáning peroxiredoxins í lungum í lungum og lungnaháþrýstingi. Þorax. 2002; 57: 157-164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, et al. Human peroxiredoxin 5 er peroxýnitrít redúktasi. FEBS Lett. 2004; 571: 161-165.
77. Holmgren A. Andoxunarefni virkni tioredoxins og glutaredoxinsystema. Andoxun Redox Signal. 2000; 2: 811-820.
78. Dickinson DA, Forman HJ. Glútaþíon í varnarmálum og merkjum: Lærdóm frá lítilli þíól. Ann NY Acad Sci. 2002; 973: 488-504.
79. Segir H. glútaþíon og hlutverk þess í frumuvirkni. Free Radic Biol Med. 1999; 27: 916-921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Samhliða þróunarsvæðin fyrir glútaþíón transferasa: uppbygging og vélbúnaður kappa ensímsins í hvatberum flokki rGSTK1-1. Lífefnafræði. 2004; 43: 52-61.
81. Robinson A, Huttley GA, Booth HS, stjórn PG. Modeling og bioinformatics rannsóknir á kappa flokki glútaþíon transferasa, spá fyrir um nýjan þriðja flutningsfamilíu með homology við prokaryotic 2-hydroxychromene-2-carboxylate isomerases. Biochem J. 2004; 379: 541-552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Algengar uppbyggingareiginleikar MAPEGda útbreiddra fjölskyldna af himnu tengdum próteinum með mjög mismunandi starfsemi í eicosanoid og glutathione umbrot. Prótein Sci. 1999; 8: 689-692.
83. Hayes JD, Pulford DJ. Glutathione S-transferasa supergen fjölskyldan: reglugerð GST og framlag ísóensímanna við krabbameinsvörn og lyfjaþol. Crit Rev Biochem Mol Biol. 1995; 30: 445-600.
84. Armstrong RN. Uppbygging, hvatakerfi og þróun glútaþíon transferasa. Chem Res Toxicol. 1997; 10: 2-18.
85. Hayes JD, McLellan LI. Glútaþíon og glútaþíon háðir ensím tákna samhliða stjórnað varnarefni gegn oxunarálagi. Free Radic Res. 1999; 31: 273-300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Uppbygging, virkni og þróun glútaþíon transferasa: áhrif til flokkunar á ófædda meðlimi fornæmis ensíms fjölskyldunnar. Biochem J. 2001; 360: 1-16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, o.fl. Glútaþíón S-transferasa Mu breyti streituvirkjunarmerkjunum með því að bæla apoptósa merki-stjórna kínasa 1. J Biol Chem. 2001; 276: 12749-12755.
88. Dorion S, Lambert H, Landry J. Virkjun p38 merkjunarferilsins með hitaáfalli felur í sér dissociation af glútaþíon S-transferasa Mu frá Ask1. J Biol Chem. 2002; 277: 30792-30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L, et al. Reglugerð um JNK-merkingu með GSTp. EMBO J. 1999; 18: 1321-1334.
90. Manevich Y, Feinstein SI, Fisher AB. Virkjun á andoxunarefninu 1-CYS peroxídoxíni krefst glútaþíónýleringa sem miðlað er með heteródimerun með pGST. Proc Natl Acad Sci US A. 2004; 101: 3780-3785.
91. Bunker VW. Hráefni, andoxunarefni og öldrun. Med Lab Sci. 1992; 49: 299-312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, et al. Almennt oxunarálag og tengsl hennar við aldur og veikindi. J er Geriatr Soc. 1996; 44: 823-827.
93. White E, Shannon JS, Patterson RE. Samband milli vítamíns og
notkun kalsíumsuppbótar og krabbamein í ristli. Krabbamein Epidemiol Biomarkers Prev. 1997; 6: 769-774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Nýjar aðferðir náttúrulegra andoxunarefnis efnasambanda í líffræðilegum kerfum: þátttaka glútaþíon og glútaþíon tengda ensím. J Nutr Biochem. 2005; 16: 577-586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Breytingar á glútatíonastöðu hjartans eftir blóðþurrð og reperfusion. Experientia. 1985; 41: 42-43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. Carotenoid róttæk efnafræði og andoxunarefni / pró-oxandi eiginleika. Arch Biochem Biophys. 2004; 430: 37-48.
97. Rice-Evans CA, Sampson J, Bramley PM, Holloway DE. Af hverju gerum við ráð fyrir að karótenóíðum verði andoxunarefni in vivo? Free Radic Res. 1997; 26: 381-398.
98. Niles RM. Signalleiðir í retínóíð kemískri meðferð og meðferð krabbameins. Mutat Res. 2004; 555: 81-96.
99. Donato LJ, Noy N. Bæling á brjóstum krabbameinsvöxtum með retínósýru: Própoptótísk gen eru markmið um retínósýru viðtaka og frumu retínósýrubindandi prótein II merki. Krabbamein Res. 2005; 65: 8193-8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, et al. Bcl-2 er lykill eftirlitsstofnanna fyrir retínósýruinnihvarfaða frumubótaðri frumudauða í taugabólga. Oncogene. 2006; 25: 5046-5055.
101. Dalton TP, Shertzer HG, Puga A. Reglugerð um genþrýsting með hvarfefnum. Ann Rev Pharmacol Toxicol. 1999; 39: 67-101.
102. Scandalios JG. Genomic viðbrögð við oxandi streitu. Í: Meyers RA, ed. Encyclopedia of Molecular Cell Biology and Molecular Medicine. Vol 5. 2nd ed. Weinheim, Þýskaland: Wiley-VCH; 2004: 489-512.
103. Ghosh R, Mitchell DL. Áhrif oxunar DNA skaða á frumefni þátttakenda á bindiefni bindingarþáttar. Nucleic Acids Res. 1999; 27: 3213-3218.
104. Marietta C, Gulam H, Brooks PJ. Einstaklingur 8, 50-sýkló-20-deoxýadenósín skaða í TATA kassi kemur í veg fyrir bindingu TATA bindiefni og dregur verulega úr umritun in vivo. DNA Repair (Amst). 2002; 1: 967-975.
105. Jackson AL, Chen R, Loeb LA. Innleiðing óstöðugleika ósatellíts
með oxandi DNA skaða. Proc Natl Acad Sci US A. 1998; 95: 12468-12473.
106. Caldecott KW. Prótein-prótín milliverkanir við DNA-brot með einangruðu DNA-broti. Biochem Soc Trans. 2003; 31: 247-251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidativ DNA skemmdir: kerfi, stökkbreytingar og sjúkdómar. FASEB J. 2003; 17: 1195-1214.
108. Jones PL, Wolffe AP. Sambönd milli krómatín samtaka og DNA metýlering við ákvörðun á gen tjáningu. Semin krabbamein Biol. 1999; 9: 339-347.
109. Girotti AW. Kerfi af peroxíðun á lípíði. J Free Radic Biol Med. 1985; 1: 87-95.
110. Siu GM, Draper HH. Umbrot malónaaldehýðs in vivo og in vitro. Fituefni. 1982; 17: 349-355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. Möguleg þátttaka lípíð-peroxíðunarafurðarins 4-hýdroxýnóníns í myndun flúrljómandi krómólípíða. Biochem J. 1986; 239: 405-409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Aldurshæfir breytingar á lípíðperoxíðhæð í lípópróteinbrotum manna sermis. J Gerontol. 1984; 39: 269-272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, o.fl. 4- Hydroxynonenal, aldehýðs vara af peroxíðmyndun himnafitu, dregur úr flutningi glutamats og hvatbera í synaptosomes. Neuroscience. 1997; 806: 85-96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Virkjun streitumerkjunarleiða með lokaafurðinni af fituprótoxíðun. 4-hýdroxý-2-nonenal er hugsanlega hvati til framleiðslu á innanfrumuperoxíði. J Biol Chem. 1999; 274: 2234-2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Virkjun EGF viðtaka með oxaðri LDL. FASEB J. 1998; 12: 665-671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hýdroxý-2-nonenal eykur framleiðslu fibrónektíns með IMR-90 lungnabólgu úr mönnum, að hluta til með virkjun á vaxtarþáttarviðtaka tengdum utanfrumuveiru p44 / 42 leið. Eiturefni Appl Pharmacol. 2002; 184: 127-135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. Útgefið 8-ísópróstan sem in vivo biomarker á oxunartruflunum á lungum hjá sjúklingum með langvinna lungnateppu og heilbrigða reykingamenn. Am J Respir Crit Care Med. 2000; 162: 1175-1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Epithelial gegndræpi, bólga og oxidant streita í loftrýmum reykinga. Am J Respir Crit Care Med. 1999; 159: 473-479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Aukið innihald tíóbárbitúrsýru hvarfefna og vetnisperoxíðs í útblástursloftþéttni sjúklinga með stöðuga langvinna lungnateppu: engin marktæk áhrif á sígarettureykingu. Respir Med. 1999; 93: 389-396.
120. Kelly FJ, Mudway IS. Prótein oxun í loft-lungum tengi. Amínósýrur. 2003; 25: 375-396.
121. Dean RT, Roberts CR, Jessup W. Fragmentation á utanfrumu og innanfrumu fjölpeptíða með sindurefnum. Prog Clin Biol Res. 1985; 180: 341-350.
122. Keck RG. Notkun t-bútýlhýdróperoxíðs sem rannsakandi fyrir metíónínoxun í próteinum. Anal Biochem. 1996; 236: 56-62.
123. Davies KJ. Prótínskemmdir og niðurbrot með súrefnishópnum. I. Almennar þættir. J Biol Chem. 1987; 262: 9895-9901.
124. Stadtman ER. Metal jón-hvataður oxun próteina: lífefnafræðilegur búnaður og líffræðileg afleiðingar. Free Radic Biol Med.
1990; 9: 315-325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Slökkt á helstu efnaskiptum ensímum með oxunarviðbrögðum í blönduðum aðgerðum: hugsanleg áhrif á veltu próteina og öldrun. Proc Natl Acad Sci US A. 1983; 80: 1521-1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oxun metíónínsleifa próteina: líffræðileg afleiðingar. Andoxun Redox Signal. 2003; 5: 577-582.
127. Stadtman ER, Levine RL. Frístilfengið oxun á frjálsum amínósýrum og amínósýruleifum í próteinum. Amínósýrur. 2003; 25: 207-218.
128. Stadtman ER. Próteinoxun í öldrun og aldurstengdum sjúkdómum. Ann NY Acad Sci. 2001; 928: 22-38.
129. Skýring E. Kvörðun og mikilvægi próteinoxunar í sýnum. Drug Metab Rev. 2000; 32: 307-326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Oxidandi streitu og frumuskemmdir. Curr Med Chem. 2004; 11: 1163-1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vascular endothelial growth factor (VEGF) og viðtaka þess. FASEB J. 1999; 13: 9-22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, o.fl. Reglugerð um hvarfefni sem mynda hvarfefni í súrefni í fibroblasts með Rac1. Biochem J. 1996; 318: 379-382.
133. Sun T, Oberley LW. Redox reglugerð um transcriptional activators. Free Radic Biol Med. 1996; 21: 335-348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redox reglugerð um DNA-bindingu c-Jun með afturkræfri S-glutathiolation. FASEB J. 1999; 13: 1481-1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. Í staðgreining á S-glútaþíónýlated prótínum eftir glutaredoxin-1 hvataða cysteine afleiður. Biochim Biophys Acta. 2006; 1760: 380-387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulation af glutaredoxin-1
tjáning í músarlíkani um ofnæmislungnasjúkdóm. Am J Respir Cell Mol Biol. 2007; 36: 147-151.
137. Filomeni G, Rotilio G, Ciriolo MR. Cell merki og glútaþíon redox kerfi. Biochem Pharmacol. 2002; 64: 1057-1064.
138. Pande V, Ramos MJ. Molecular recognition of 15-deoxydelta (12,14) prostaglandín J (2) með kjarnaþætti-kappa B og öðrum frumuprótínum. Bioorg Med Chem Lett. 2005; 15: 4057-4063.
139. Perkins ND. Sameining klefi-merkja leið með NF-kappaB og IKK virka. Nat Rev Mol Cell Biol. 2007; 8: 49-62.
140. Gilmore TD. Kynning á NF-kappaB: leikmenn, leiðir, sjónarmið. Oncogene. 2006; 25: 6680-6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Einstaka hlutverk tioredoxins í frumum og kjarna. Tvær skref kerfi af redox reglugerð um uppskrift þáttur NF-kappaB. J Biol Chem. 1999; 274: 27891-27897.
142. Ward PA. Hlutverk viðbótar, chemokines og reglulegrar cýtókína við bráða lungaskaða. Ann NY Acad Sci. 1996; 796: 104-112.
143. Akira S, Kishimoto A. NF-IL6 og NF-kB í frumuskiptingu frumna. Adv Immunol. 1997; 65: 1-46.
144. Meyer M, Schreck R, Baeuerle PA. H2O2 og andoxunarefni hafa gagnstæða áhrif á virkjun NF-kappa B og AP-1 í ósnortnum frumum: AP-1 sem afleidd andoxunarefnahrif. EMBO J. 1993; 12: 2005-2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Redox reglugerðin um fos og Jun DNA bindandi virkni in vitro. Vísindi. 1990; 249: 1157-1161.
146. Galter D, Mihm S, Droge W. Einstök áhrif glútaþíón tvísúlfíðs á kjarnorkuþrýstingsþáttum kB og virkjunarprótín-1. Eur J Biochem. 1994; 221: 639-648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. AP-1 transkriptunarvirkni er stjórnað með beinni tengslum milli thioredoxins og Ref-1. Proc Natl Acad Sci US A. 1997; 94: 3633-3638.
Post Fyrirvari
Upplýsingarnar hér á „El Paso, TX Oxidative Stress og andoxunarefni vörn" er ekki ætlað að koma í stað einstaklingssambands við hæfan heilbrigðisstarfsmann eða löggiltan lækni og er ekki læknisráðgjöf. Við hvetjum þig til að taka heilbrigðisákvarðanir byggðar á rannsóknum þínum og samstarfi við hæfan heilbrigðisstarfsmann.
Blogg upplýsingar og umfang umræður
Upplýsingar umfang okkar takmarkast við kírópraktík, stoðkerfi, nálastungumeðferð, líkamleg lyf, vellíðan, stuðla að orsök truflanir í innyflum innan klínískra kynninga, tilheyrandi sematovisceral reflex klínískt gangverki, subluxation complexes, viðkvæm heilsufarsvandamál og/eða greinar, efni og umræður um hagnýt lyf.
Við veitum og kynnum klínískt samstarf með sérfræðingum úr ýmsum greinum. Sérhver sérfræðingur er stjórnað af faglegu starfssviði sínu og lögsögu sinni í leyfisveitingu. Við notum hagnýtar heilsu- og vellíðunarreglur til að meðhöndla og styðja umönnun vegna meiðsla eða sjúkdóma í stoðkerfi.
Vídeó okkar, færslur, efni, viðfangsefni og innsýn ná yfir klínísk atriði, málefni og efni sem tengjast og styðja beint eða óbeint klínískt umfang okkar starfs.*
Skrifstofa okkar hefur sanngjarnt reynt að veita stuðningstilvitnanir og hefur bent á viðeigandi rannsóknarrannsóknir sem styðja innlegg okkar. Við bjóðum afrit af stuðningsrannsóknum sem eru tiltækar eftirlitsnefndum og almenningi sé þess óskað.
Við skiljum að við fjöllum um mál sem krefjast frekari skýringa á því hvernig það getur hjálpað til við ákveðna umönnunaráætlun eða meðferðarreglur; vinsamlegast ekki hika við að ræða um efnið hér að ofan Dr. Alex Jimenez, DC, Eða hafðu samband við okkur á 915-850-0900.
Við erum hér til að hjálpa þér og fjölskyldu þinni.
Blessun
Dr. Alex Jimenez D.C., MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
netfang: coach@elpasofunctionalmedicine.com
Með leyfi sem Doctor of Chiropractic (DC) í Texas & Nýja Mexíkó*
Texas DC leyfisnúmer TX5807, New Mexico DC Leyfisnúmer NM-DC2182
Með leyfi sem hjúkrunarfræðingur (RN*) í florida
Flórída leyfi RN leyfi # RN9617241 (Stjórn nr. 3558029)
Samningsstaða: Fjölríkis leyfi: Leyfi til að æfa í 40 ríki*
Stúdentspróf: ICHS: MSN* FNP (Fjölskylduhjúkrunarfræðingaáætlun)
Dr. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Stafræna nafnspjaldið mitt
 Aftur tökum við vel á móti þér¸
Aftur tökum við vel á móti þér¸
